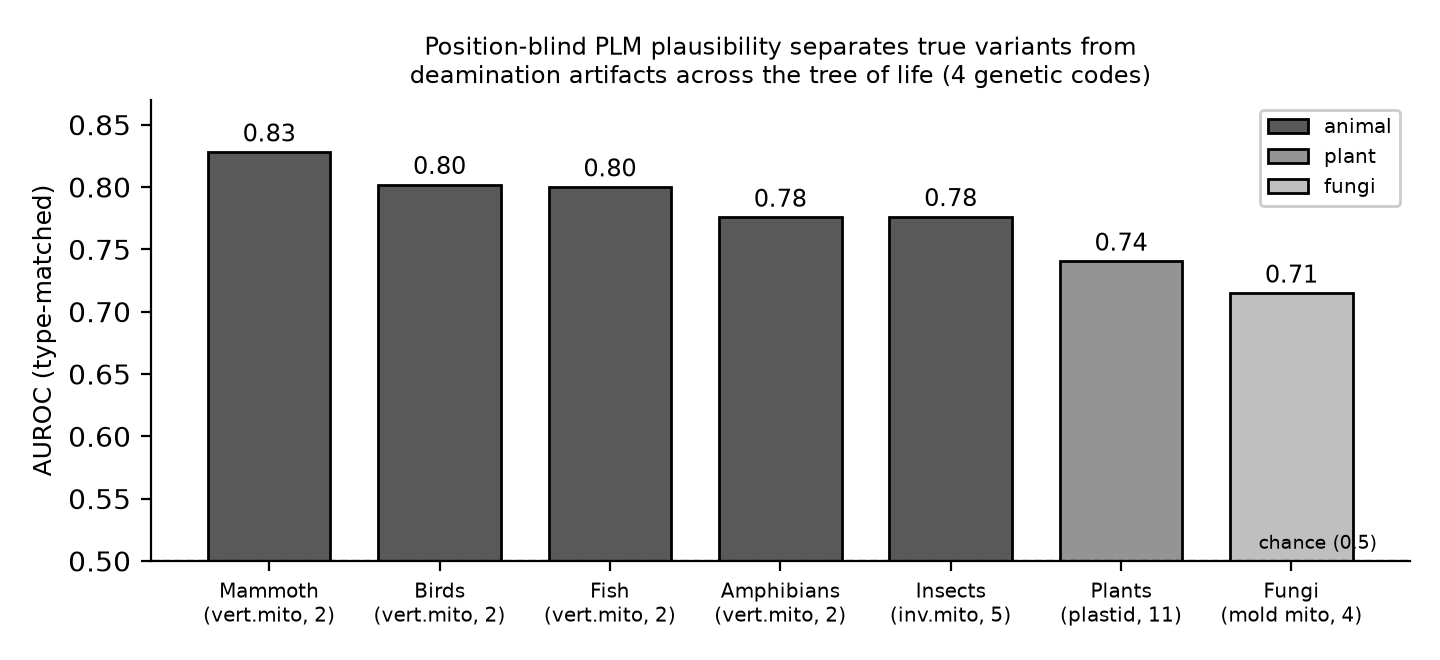
マンモスだけの現象ではありません。4つの遺伝暗号にまたがる7クレード(脊椎動物ミト・無脊椎ミト・色素体・糸状菌ミト)すべてで、位置ブラインドのPLM妥当性は真の変異と脱アミノ化損傷を有意に判別しました。
- 研究テーマ
- 古代DNAの真贋判定 × タンパク質言語モデル(読み取り位置に依らない真正性シグナル)
- 著者
- Masato Suzuki(Masafy)/ORCID 0009-0000-7977-2756
- モデル
- ESM-2(650M, esm2_t33_650M_UR50D)/wild-type marginal による対数尤度比
- データ
- NCBI RefSeq のミトゲノム・色素体ゲノム+ENAのマンモス生リード(ADYCHA, 約120万年前を含む)
- 公開先
- GitHub(public) / Zenodo(DOI):論文PDF(日英)+再現コード
- 関連特許
- 特願2026-173418(PLM-aDNA真正性判定方法/システム/プログラム、出願係属中)
- ライセンス
- CC BY 4.0(論文・データ)
※ 本ページは著者本人が自らの判断で公開している研究情報です。特許は出願済みのため公開に支障はありません(識別番号・住所等の管理情報は非掲載)。
「どこで読まれたか」ではなく、
「配列としてありえるか」で見抜く。
古代DNAの真贋を疑うとき、これまでの主役は読み取り末端の損傷パターンでした。断片の端でC→T/G→Aが増えるという、位置に依存した特徴です。強力ですが、断片が短い・被覆が浅い・そもそも「この1塩基の置換は本物か損傷か」を問いたい、といった場面では届きません。
この研究の着眼はシンプルです。タンパク質は、意味のある配列しか生き残らない。だから、ある置換がタンパク質として「ありえる」ものかを大規模タンパク質言語モデルに尋ねれば、本物の進化的変異は妥当なまま・脱アミノ化損傷は妥当性を落とす——読み取り位置の情報を一切使わずに、両者を分けられるはずだ、と。実際にそれが成り立つことを、マンモスから生命樹全域まで確かめました。
① 配列の“もっともらしさ”
ESM-2 が測るタンパク質としての妥当性を、置換前後の対数尤度比でスコア化。
② 位置ブラインド
読み取り位置を一切使わない、配列そのものからの独立シグナル。ここが核心。
③ 生命樹で普遍
動物・植物・菌、4つの遺伝暗号を跨いで一貫して成立する。
損傷と変異は、なぜ紛らわしいのか。
脱アミノ化は、時間とともにシトシンをウラシル(読み取りではT)に変える化学反応。結果として現れる C→T / G→A は、本物の突然変異と塩基レベルで区別がつかない。
既存の真贋判定(mapDamage 系)は、断片末端に損傷が集中する「位置パターン」に依存する。読み取り構造がある集団では強力だが、位置情報が乏しい/単一サイトを問いたい場面では弱い。
つまり必要なのは、読み取り位置に依らない、配列そのものから来る独立した真正性シグナル。位置ベースの手法と相補的に効くものが欲しい。
実データで、順を追って。
※ すべて論文の実測図(グレースケール)。GitHub / Zenodo で再現可能。
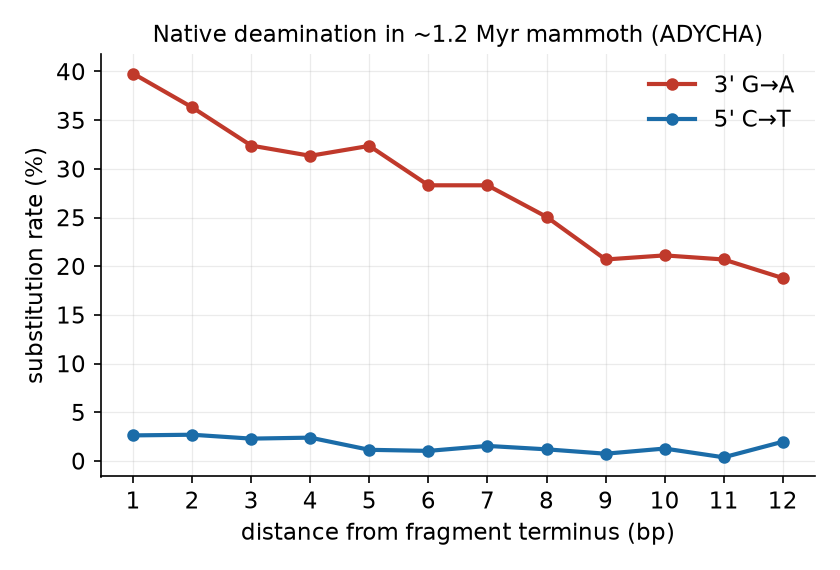
約120万年前のマンモス(ADYCHA)に刻まれた脱アミノ化。断片末端で置換率が跳ね上がる——これが古代DNAの“指紋”であり、位置ベース手法が使ってきた特徴です。
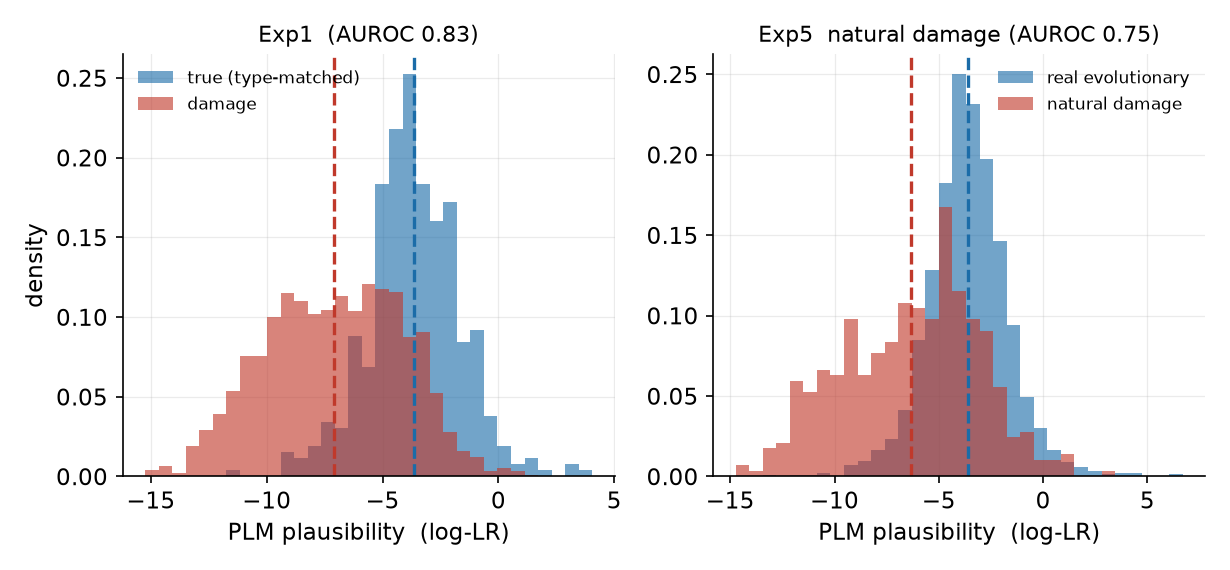
ESM-2 の妥当性スコア(対数尤度比)の分布。真の変異(青)は高く、損傷(赤)は低い側に寄り、位置情報を使わずとも両者が分かれます。
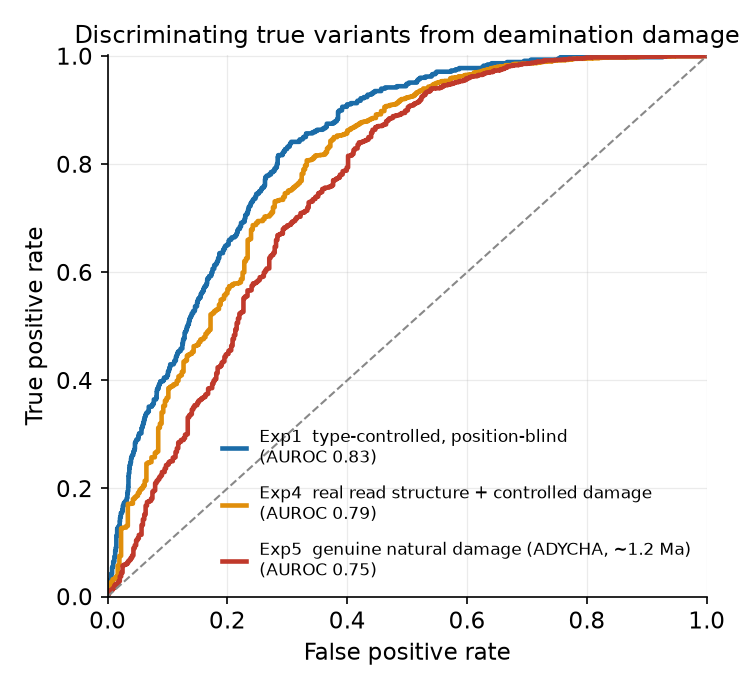
理想条件から実データまで3つの設定で AUROC 0.75〜0.83。約120万年前の天然の損傷でも 0.75 を保ちます。
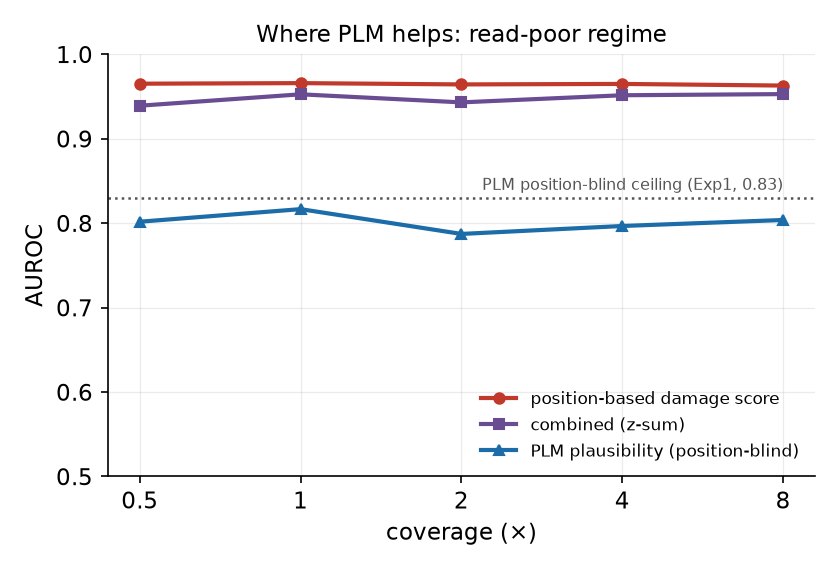
読み取り構造がそろえば位置ベース手法は非常に強い。PLMの価値は、読み取り位置に依らず・被覆に左右されずに効く独立シグナルである点にあります(相補的)。
そして本命。マンモス固有の芸当ではありません。脊椎動物ミト・無脊椎ミト・色素体・糸状菌ミトの4暗号・7クレードすべてで、位置ブラインドPLMは損傷と真の変異を有意に判別しました。
7クレード、すべてで有意。
型統制・位置ブラインドの ESM-2 妥当性による AUROC。すべてのクレードで、真の変異は損傷より有意に妥当(Mann–Whitney)。
| クレード | 系統 | 遺伝暗号 | AUROC(型統制) | p 値 |
|---|---|---|---|---|
| マンモス | 動物 | 脊椎動物ミト(2) | 0.828 | ~1×10⁻¹⁰⁵ |
| 鳥類 | 動物 | 脊椎動物ミト(2) | 0.802 | 1.8×10⁻⁷⁴ |
| 魚類 | 動物 | 脊椎動物ミト(2) | 0.800 | 6.0×10⁻⁷¹ |
| 両生類 | 動物 | 脊椎動物ミト(2) | 0.776 | 3.2×10⁻⁶⁸ |
| 昆虫 | 動物 | 無脊椎動物ミト(5) | 0.776 | 4.1×10⁻⁷⁹ |
| 植物 | 植物 | 色素体(11) | 0.740 | 2.4×10⁻¹⁵⁹ |
| 菌類 | 菌 | 糸状菌ミト(4) | 0.715 | 1.3×10⁻⁴⁷ |
勾配は 動物(0.78〜0.83)> 植物(0.74)> 菌(0.72)。菌がやや弱いのは、ミトコンドリアORFの混入・遺伝子命名の揺れ・分岐の大きさによると考えられます。
この研究が示したこと。
位置に依らない真正性シグナル
読み取り末端の損傷パターンに頼らず、配列そのものから真贋の手がかりを得られる。位置ベース手法と相補的。
既存モデルの転用
古代DNA向けに新しく学習させるのではなく、汎用のタンパク質言語モデルをそのまま使える。追加学習なしで機能する。
生命樹全域で普遍
動物・植物・菌、4つの遺伝暗号を跨いで成立。特定の生物に限らない、一般的な性質であることを示した。
自宅GPUと、公開データだけで。
実験は、ENA から落としたマンモスの生リードと、NCBI RefSeq の公開ゲノム、そして自前の計算環境で完結しています。ESM-2 は一般公開のモデルをそのまま利用。マンモスで確かめた結果を、鳥・魚・両生・昆虫・植物・菌へと1時間ほどで生命樹全域に広げ、すべてで再現しました。
論文(日英)・再現コード・図版はすべて GitHub と Zenodo(DOI) で公開。関連する発明は 特許(特願2026-173418)として出願済みです。